За да се говори за двадесетте аминокиселини, които изграждат протеиновите структури и модифицираните, би било необходимо да се опишат поне дванадесет специализирани метаболитни пътища.
Но защо клетките използват толкова много метаболитни пътища, които изискват енергия (например за регенериране на каталитичните места на ензимите), всяка от които с ензимно наследство, за катаболизиране на аминокиселини? От почти всички аминокиселини е възможно да се получат чрез специализирани пътища метаболити, които в малка част се използват за производство на енергия (например чрез глюконеогенеза и пътя на кетонните тела), но които преди всичко водят до образуването на комплекс молекули, с голям брой въглеродни атоми (например от фенилаланин и тирозин, хормоните се произвеждат в надбъбречните жлези, които са специализирани за тази цел); ако от една страна би било лесно да се произвежда енергия от аминокиселини, от друга би било сложно изграждането на сложни молекули, започвайки от малки молекули: катаболизмът на аминокиселините им позволява да експлоатират скелета си, за да получат по -големи видове.
Две или три хектограми на аминокиселини се разграждат ежедневно от здрав индивид: 60-100 g от тях произлизат от протеините, въведени с храната, но над 2 хектограми се получават от нормалния оборот на протеини, които са неразделна част от организма ( аминокиселините на тези протеини, които са повредени от окислително -възстановителни процеси, се заменят с други и се катаболизират).
Аминокиселините дават енергиен принос по отношение на АТФ: след отстраняване на а-амино групата, останалият въглероден скелет от аминокиселини, след подходящи трансформации, може да влезе в цикъла на Кребс. Освен това, когато доставките на хранителни вещества липсват и количеството глюкоза намалява, се активира глюконеогенезата: за глюконеогенетични аминокиселини се казва, че тези, които след подходящи модификации могат да бъдат въведени в глюконеогенезата; глюконеогенетичните аминокиселини са тези, които могат да се превърнат в пируват или във фумарат (фумаратът може да се превърне в малат, който напуска митохондриите и в цитоплазмата се трансформира в оксалоацетат, от който може да се получи фосфоенол пируват). оцет-ацетат.
Току -що описаният е много важен аспект, тъй като аминокиселините могат да поправят недостиг на захар в случай на незабавно гладуване; ако гладуването продължава, след два дни се намесва липидният метаболизъм (тъй като протеиновите структури не могат да бъдат атакувани твърде много), именно в тази фаза, тъй като глюконеогенезата е много ограничена, мастните киселини се превръщат в ацетил коензим А и кетонни тела. От по -нататъшно гладуване мозъкът също се адаптира да използва кетонните тела.
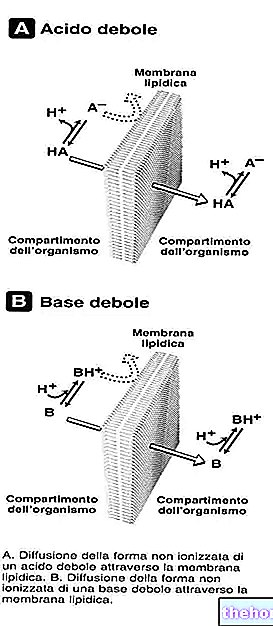
Прехвърлянето на а-амино групата от аминокиселини става чрез реакция на трансаминиране; ензимите, които катализират тази реакция, казват, всъщност, трансаминази (или амино трансфераза). Тези ензими използват ензимен кофактор, наречен пиридоксал фосфат, който се намесва с неговата алдехидна група. Пиридоксал фосфатът е продукт на фосфорилирането на пиридоксин, който е витамин (В6), който се намира главно в зеленчуците.
Трансаминазите имат следните свойства:
Висока специфичност за а-двойка кетоглутарат-глутамат;
Те са кръстени на втората двойка.
Ензимите на трансаминазите винаги включват двойката α кетоглутарат-глутамат и се разграничават според втората включена двойка.
Примери:
L "аспартат трансаминаза т.е. GOT (глутамат-осал ацетат трансаминаза): ензимът прехвърля а-амино групата от аспартат в а-кетоглутарат, получавайки оксалоацетат и глутамат.
L "аланин трансаминаза т.е. GTP (глутамат-пируват трансаминаза): ензимът прехвърля а-амино групата от "аланин в" а-кетоглутарат, получавайки пируват и глутамат.
Различните трансаминази използват α-кетоглурат като акцептор на аминогрупата на аминокиселините и я превръщат в глутамат; докато образуваните аминокиселини се използват по пътя на кетонните тела.
Този тип реакция може да се случи в двете посоки, тъй като те се разрушават и образуват връзки със същото енергийно съдържание.
Трансаминазите са както в цитоплазмата, така и в митохондрията (те са предимно активни в цитоплазмата) и се различават по своята изоелектрична точка.
Трансаминазите също са способни да декарбоксилират аминокиселини.
Ще трябва да има начин да се превърне глутамат обратно в α-кетоглутарат: това става чрез дезаминиране.
Там глутамат дехидрогеназа това е ензим, способен да трансформира глутамат в α-кетоглутарат и следователно да превръща аминогрупите на аминокиселините, намиращи се под формата на глутамат, в амоняк. Това, което се случва, е редокс процес, който преминава през междинния α-амино глутарат: амонякът и α-кетоглутаратът се освобождават и се връщат в кръвообращението.
След това изхвърлянето на аминогрупите на аминокиселините преминава през трансаминазите (които се различават в зависимост от субстрата) и глутамат дехидрогеназата, която определя образуването на амоняк.
Има два вида глутамат дехидрогеназа: цитоплазмена и митохондриална; кофакторът, който също е косубстрат на този ензим е NAD (P) +: глутамат дехидрогеназата използва или NAD +, или NADP + като акцептор на редуцираща мощност. Цитоплазмената форма предпочита, макар и не изключително, NADP +, докато митохондриалната форма предпочита NAD +. Митохондриалната форма има за цел да изхвърли амино групи: води до образуване на амоняк (който е субстрат за специализиран ензим в митохондрията) и NADH (който се изпраща към дихателната верига). Цитоплазмената форма работи в обратна посока, т.е. използва амоняк и α-кетоглутарат, за да даде глутамат (който има биосинтетична дестинация): тази реакция е редукционна биосинтеза и използваният кофактор е NADPH.
Глутамат дехидрогеназата действа, когато е необходимо да се изхвърлят аминогрупите от аминокиселини като амоняк (чрез урина) или когато скелетите на аминокиселините са необходими за производство на енергия: следователно този ензим ще има като отрицателни модулатори системите, които са индикация с добра енергийна наличност (ATP, GTP и NAD (P) H) и като положителни модулатори, системите, които показват нужда от енергия (AMP, ADP, GDP, NAD (P) +, аминокиселини и хормони на щитовидната жлеза).
Аминокиселините (главно левцин) са положителни модулатори на глутамат дехидрогеназата: ако аминокиселините присъстват в цитоплазмата, те могат да се използват за синтез на протеини или трябва да се изхвърлят, защото не могат да се натрупват (това обяснява защо аминокиселините са положителни модулатори ).
Изхвърляне на амоняк: цикъл на карбамид
Рибите изхвърлят амоняка, като го въвеждат във вода през хрилете; птиците го превръщат в пикочна киселина (която е продукт на кондензация) и го елиминират с изпражненията. Да видим какво се случва при хората: казахме, че глутамат дехидрогеназата превръща глутамата в α- кетоглутарат и амоняк, но не сме казали, че това се случва само в митохондриите на черния дроб.
Фундаментална роля при изхвърлянето на амоняк чрез цикъла на карбамида играят митохондриалните трансаминази.

Въглеродният диоксид, под формата на бикарбонатен йон (HCO3-), се активира от биотиновия кофактор, образувайки карбоксибиотин, който реагира с амоняк, за да даде карбаминова киселина; следващата реакция използва АТФ за прехвърляне на фосфат върху карбаминовата киселина, образувайки карбамил фосфат и ADP (превръщането на АТФ в АДФ е движещата сила за получаване на карбоксибиотин). Тази фаза се катализира от карбамил фосфат синтетаза и се среща в митохондриите. Карбамил фосфат и орнитин са субстрати за ензима орнитин транс карбамилаза което ги превръща в цитрулин; тази реакция протича в митохондриите (хепатоцитите). Произведеният цитрулин напуска митохондриите и в цитоплазмата преминава под "действието на"аргинин сукцинат синтетаза: има сливане между въглеродния скелет на цитрулин и този на аспартат чрез нуклеофилна атака и последващо елиминиране на водата. Ензимът аргинин сукцинат синтетаза изисква молекула АТФ, така че има енергийно свързване: хидролизата на АТФ до АМР и пирофосфат (последният след това се превръща в две молекули ортофосфат) става чрез изтласкване на молекула d "вода от субстрата а не чрез действието на водата на средата.
"Следващият ензим е"аргинин сукциназа: този ензим е в състояние да раздели аргинин сукцинат на аргинин и фумарат в цитоплазмата.
Цикълът на карбамид се завършва от ензима аргиназа: получават се урея и орнитин; уреята се изхвърля от бъбреците (урина), докато орнитинът се връща в митохондрията и възобновява цикъла.
Цикълът на урея е обект на непряка модулация от аргинин: натрупването на аргинин показва, че цикълът на урея трябва да се ускори; модулирането на аргинин е непряко, тъй като аргининът модулира положително ензима ацетил глутамат синтетаза. Последният е в състояние да прехвърли ацетилова група върху азота на глутамат: Образува се N-ацетил глутамат, който е директен модулатор на ензима карбамил-фосфо синтетаза.
Аргининът се натрупва като метаболит на урейния цикъл, ако производството на карбамил фосфат не е достатъчно за изхвърляне на орнитина.
Урея се произвежда само в черния дроб, но има и други места, където се извършват първоначалните реакции.
Мозъкът и мускулите използват специални стратегии за елиминиране на амино групи. Мозъкът използва много ефективен метод, при който се използва ензим глутамин синтетаза и ензим глутамаза: първият присъства в невроните, докато вторият се намира в черния дроб. Този механизъм е много ефективен по две причини:
Две амино групи се транспортират от мозъка до черния дроб в един -единствен носител;
Глутаминът е много по -малко токсичен от глутамата (глутаматът също осъществява трансфер на неврони и не трябва да надвишава физиологичната концентрация).
При рибите подобен механизъм пренася аминогрупата от аминокиселини в хрилете.
От мускула (скелетния и сърдечния) аминогрупите достигат черния дроб чрез глюкозно-аланиновия цикъл; участващият ензим е глутамин-пируват трансаминазата: позволява транспонирането на амино групи (които са под формата на глутамат), превръщането на пируват в аланин и в същото време глутамат в а-кетоглутарат в мускула и катализиране на обратен процес в черния дроб.
Трансаминазите с различни задачи или позиции също имат структурни различия и се определят чрез електрофореза (те имат различни изоелектрични точки).
Наличието на трансаминази в кръвта може да бъде симптом на увреждане на черния дроб или сърцето (т.е. увреждане на тъканите на чернодробните или сърдечните клетки); трансаминазите са в много високи концентрации както в черния дроб, така и в сърцето: чрез електрофореза е възможно да се установи дали увреждането е настъпило в черния дроб или сърдечните клетки.